Bigfoot Encounters
POSSIBLE ANCESTRY OF SASQUATCH AND ITS EURASIAN KIN
By Charles A. Reed
Lacking definitive physical remains of the Sasquatch, we can only speculate on its affinities. Here, Charles Reed engages in a rather exhaustive coverage of all the reasonable possibilities and gives many of the pros and cons for each of them. His analysis not only deals with the anatomical and behavioral data for Sasquatch and the various candidates, but also handles geographical and climatological arguments as well. His conclusion is that Gigantopithecus is the most likely candidate, but also stresses "this is far from certain." In addition, he notes the possibility that more than one kind of unknown hominoid may be involved in the reports--especially in Eurasia. Ivan Sanderson long ago suggested that many different kinds of unknown hominoids existed, but few have followed this lead until now.
That Science could have missed one bipedal primate is a revolutionary thought. That Science has missed two or more of them, at first thought borders on the absurd. But one must realize that what ever procedures are responsible for missing the first, these would necessarily apply to any and all others that might exist. Professor Reed is a cautious scholar, neither an enthusiast nor a skeptic, who has brought a broad range of evidence to bear on the diversity of opinions about Sasquatch.
Note at once that
we are not considering a possible native of North America, only; if Sasquatch
exists; it is a population whose presence in Eurasia is at least as definite
as in North America. This situation demands due consideration of paleo-geographical
evidence as well as that of morphology and possible evolution, No matter
where the ancestors of Sasquatch first evolved, that clan has spread out
to be present on both sides of the northern Pacific. Sometime, probably
when they had evolved to be nearly similar to the present Sasquatch (remembering
always, if there is a Sasquatch) extension of range had occurred across
Bering Strait, when that strait was instead an isthmus, as it was at different
times during the Cenozoic. This latter paleo-geographic factor, usually
ignored by hominologists, allows us to consider periods other than the
Pleistocene for movement of sasquatches or their ancestors 'from one continent
to the next. However, even if movement of a population of primates from
Alaska to Siberia or vice versa did not occur during a glacial period
of the Pleistocene, that northern route would have been cold temperate
at any time after the Eocene, whereas by contrast most primates during
the last 30,000,000 years have ranged and do range climatically from the
warm tropics to warm temperate regions. Even those few nonhuman primates,
all monkeys, which do live in areas with winter snows do not penetrate
the north-temperate forests of the high latitudes, as Sasquatches or their
ancestors must have done.
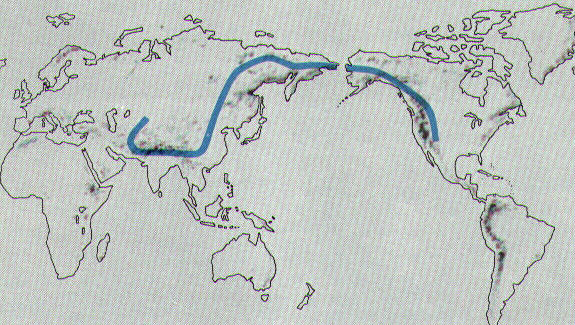
Thus the northern
transcontinental Bering Isthmus was a cold region, not necessarily "Arctic" as it was during each glacial phase of the Pleistocene, but yet cold.
Also, the passage across it of any population was necessarily a slow one;
a population of animals doesn't rush across a new area but instead spreads
slowly, probably due to increase in population pushing a few individuals
per generation a few hundred meters or so further into new territory.
The animals making that crossing, as true of all which did so after the
Eocene, had to be sufficiently adapted anatomically, physiologically,
and behaviorally to survive the cold, the the wet, and the wind; lacking
the technology which humans use and have used in the far north, heavy
pelage and layers of fat were undoubtedly the major factors of that adaptation.
Too, lacking a ruminant's stomach and probably also lacking the less efficient
but still useful large caecum of some other ungulates, being omnivorous
would have been useful, as would also the energy-conserving practice of
daily or even weekly periods of winter sleep--but of these possibilities
we know nothing.
POTENTIAL ANCESTRAL
GROUPS
I. Western Hemisphere:
1. Plesiadapiformes, the most primitive suborder of Primates, had its major adaptive radiation in the Paleocene of North America and Europe. In Europe only one genus lived into the earliest Eocene; but of the four or five North American families known, two persisted into the mid-Eocene and one into the latest Eocene. Of the several genera, most were the size of shrews and mice, with one as large as a squirrel. Mostly they had extremely specialized dentitions, with large, rodent-like incisors, loss of canines, and variable numbers of premolars, which specialization's preclude them from the possibility of being ancestral to later primates.
2. The "true" primates (with grasping hands and feet and various other indisputable characters of our order) appeared suddenly in considerable profusion in North America and Europe and in the earliest Eocene, some 55,000,000 years ago. They have been complexly (and bewilderingly) classified and reclassified in different combinations, but from these earliest "true" primates all living ones have descended, although the pattern of that descent is often not now known. Thinking at present only of North America, three families are usually recognized (Adapidae, Anaptomorphidae, and Omomyidae) or sometimes often two, uniting the anaptomorphids and omomyids into a single family. All of these North American primates, with a single exception, seem to have become extinct in the latest Eocene and earliest Oligocene. The exception is a genus of omomyid with the unpronounceable name of Ekgmowechashala, of which a few jaws have been found in the early Miocene of South Dakota. As no fossil of a primate has been found in North America from the early Oligocene to the early Miocene, a period of approximately 11,000,000 years, this situation clearly illustrates the principle of the incompleteness of the fossil record, since Ekgmowechashala certainly had living ancestors during that period of 11,000,000 years.
Even with the above lesson of a known major gap in the fossil record of North American primates, I discount the probability of a continuing but unknown population of Miocene and post-Miocene primates on North America, of which living representatives would be giant bipeds looking and acting more like hominoids (apes and hominids) than like prosimians. The time between Ekgmowechashala and the present was nearly 25,000,000 years, and indeed during that same period of 25,000,000 years in the Old World one lineage of primitive pongids did evolve into the gibbons, the Great Apes, and the hominids.
Thus, I don't think that Sasquatch and his modern relatives (presuming, as always that Sasquatches exist) underwent their evolution on North America, in spite of the numerous primates living on that continent from the early Paleocene and through the Eocene and surviving, indeed, into the Miocene.
3. Platyrrhini (flat-nosed primates): Primates of unknown ancestry had somehow reached South America from either Africa or North America, and thus across a considerable water gap in either case, by the early Oligocene, or possibly even earlier. South America some 30,000,000 years ago was an island continent, separated from North America and drifting slowly westward, away from Africa. During the long period of subsequent isolation, the South American primates (Platyrrhini) underwent a remarkable and remarkably interesting adaptive radiation, different lineages evolving to become fitted to different ecological niches in the vast forests, which for a considerable time extended south almost to the Straits of Magellan. However, the platyrrhines remained arboreal, even though a few kinds do sometimes forage or move between isolated clumps of trees on the ground. All of the varied types of South American primates are recognizable as "monkeys" to the man and child at the zoo; of these types, the marmosets are similar in size and form to many of the prosimians of the Old World, the majority of platyrrhines are more similar to the general types of monkeys of Asia and Africa, and one kind, the spider monkey, is superficially similar in size and proportions to the gibbon, the aerialist of southeastern Asia, but the spider monkey is a less versatile acrobat. All South American monkeys have tails, but, contrary to popular opinion, most of them don't have prehensile tails.
When South and North America became united at the Isthmus of Panama some four million years ago, a few kinds of these platyrrhine monkeys spread north through the continuous tropical jungle to its borders in southern Mexico, and one can imagine one such species, more ground-adapted than the others, continuing that northward extension of range, evolving large size and more efficient bipedalism in the process, but losing its tail. We do have a model for such a possibility; the howler monkey, at a maximum of 40 pounds, is the largest platyrrhine and (aside from the laryngeal specializations) could provide an ancestral type if it became more ground-living at forest-fringes, and became bipedal in so doing. Some of the larger platyrrhines do indeed, become bipedal when on the ground or large horizontal branches, and some of medium-size as the capuchin, easily acquire bipedalism when trained to beg with cup in hand. Increasing size and occasional bipedality, thus, are characters already present in South American monkeys, and natural selection has often acted upon no stronger characters to produce new variant populations in several million years.
Would such a giant bipedal monkey still be recognizable as a monkey, or would it--as all observers of Sasquatch have agreed --appear to be a large ape or human? And would it, if so evolved and spread northward, have continued across a Bering Bridge to furnish an Asiatic population, as attested by Russian and Chinese observers? Would 4,000,000 years have been sufficient for such an evolution?
On all counts, I think not; I believe that our Sasquatch is (if existing) a "hominoid" (member of the Super family Hominoidea, pongids plus hominids).
II. Eastern Hemisphere
All of the living non-prosimian Old World primates are included in the
Catarrhini, further subdivided into Cercopithecoidea (monkeys) and Hominoidea
(apes and hominids).
1. Old World Monkeys: The members of this group, in my opinion, can have little more relationship to our hypothetical Sasquatch than do the South American monkeys, and so, considering the very hominoid appearance reported by all viewers of Sasquatch and its Old World relatives, I shall give no consideration in this survey of cercopithecoids as possible ancestral Sasquatches.
2. Hominoidea: All of the fossil and living populations included in these groups are so similar that they would be placed in the same family if it were not that modern man for historical reasons based on his own ego segregates the apes and their ancestors into a separate family, the Pongidae. Even so, a few taxonomists at present have made the preliminary step of including gorillas and chimpanzees in the Hominidae. I shall not, for these and other reasons, attempt in this survey to find that supposed line between ourselves and our closer relatives, but shall call us all simply hominoids.
A. Gigantopithecus: Most persons who have considered the problem of the ancestor for Sasquatch have suggested this large-jawed, large-toothed hominoid of the Plio-Pleistocene of India and China as the most probable candidate. In spite of a voluminous literature on the supposed structure, behavior, environment, diet, adaptations, phylogeny, etc., of this candidate, we really know remarkably little about Gigantopithecus. The recovered fossils consist of only four broken jaws assigned to two species and some 1300 whole or broken teeth of one of the species. Mostly it has been called a giant ape belonging to the Mio-Pliocene group called the dryopithecines, but a few investigators, using the same data of course, have labeled it as a primitive hominid, which has become highly specialized in its own way (Eckhardt 1975). Without further fossils or a living representative we will never know!
The known range in time and space of Gigantopithecus is from about 6,000,000 years ago in northwestern India (one broken mandible named G. bilaspurensis) to approximately 500,000 years ago in south-central China ((7. blacki), three broken mandibles and approximately 1300 teeth or parts thereof (Rukang 1982a). In spite of the relatively few fossils of few bodily parts, the populations belonging to Gigantopithecus certainly lived earlier and later than the times known for the fossils, and also over a much wider area (possibly most of Asia or beyond) than the regions of the few known fossils (Rukang 1982b).
I wish to stress how little we really know about Gigantopithecus; we know its dentition and that it wore its teeth fiat; we know that it didn't have typically apish projecting canines, and that sometimes, as with robust australopithecines, the canines were worn fiat to become a part of the grinding plane of the cheek teeth; we know it had the largest jaws and teeth of all primates; we know that the known fossils were found in tropical and/or semitropical forested environments; and we know that 8.9% of the recovered teeth of G. blacki had caries, suggesting for that species a diet high in carbohydrates (this factor, plus the wear on the cheek-teeth, has led some investigators to suggest that the diet, like that of the panda, emphasized bamboo). We don't know the size of individuals of either species of Gigantopithecus; size of jaw is only a rough measure of the size of the animal possessing the jaw and may indicate more about the roughage in the diet and the power and size of the masticator muscles than height and weight of the animal. Gigantopithecus was obviously a large primate, but whether larger or smaller than a gorilla we don't know. We don't know if it could climb or not, but as true of even large gorillas at present, some climbing presumably was both possible and practiced. Above all, we don't know whether Gigantopithecus was typically quadrupedal or bipedal.
When suggesting Gigantopithecus as an ancestor, or ancestral type for Sasquatch, most investigators obviously have in mind the mid-Pleistocene G. blacki from China as being closer chronologically and geographically to the North American present. However, on simple climatological bases, I will argue for the earlier G. bilaspurensis of India or a contemporaneous but unknown species elsewhere in Asia as a more reasonable ancestral possibility. The finding of the single jaw of this species in northwestern India does not mean that the total population lived only there, or lived only in the semi-tropical and/or tropical forests of that area; like pumas, tigers, and macaques (to mention a few mammals among many), G. bilaspurensis or other gigantopithecines may have had wide temperature tolerances while yet it was limited to more or less open forests. Such open forest, at least in northeastern Siberia during the early and middle Pliocene, stretched to the Arctic ocean, and the annual mean temperature, as based upon the known plants of the area at the time (conifers such as spruce, pine, hemlock, larch, and fir, but with a considerable mixture of deciduous angiosperms as oak, birch, elder, hornbeam, and elm), was more equable and not as cold as at present (Petrov 1967). In other words, the environment of northeastern Siberia was quite similar to that of parts of British Columbia and southern Alaska where Sasquatches are reputed to be living today, We can imagine thus, but cannot now prove, that the mid-Pliocene population of G. bilaspurensis, represented only by a partial mandible in India, could also have been living throughout much of the forested areas of Asia, and that the more northern individuals of that population would have become adapted by factors of specialized behavior, denser pelage, and greater fat deposits to colder northern environments, probably to live quite comfortably there as sasquatches in British Columbia seem to do today.
I do not credit any
population of Gigantopithecus with the ability to construct boats or rafts,
or even to survive drifting on a log any distance in the open sea, but
no such conjectures are necessary; a Bering Isthmus was present during
the early and middle Pliocene, and the local environment on that isthmus "...may have been a mixture of muskeg vegetation and woodlands dominated
by birch, aspen, alder and willow" (Hopkins 1967).
In other words, the environment across the strait was very similar to
that of the Siberian corner to which the Sasquatches were possibly already
adapted and, spreading slowly generation by generation, they would never
have known that they had in time penetrated Alaska and thus another continent.
On the Alaskan side,
the new intruders would have found a continuation of the same type of
environment in which they may have been living for several hundreds of
thousands of years (Hopkins et al. 1971; Wolfe 1972).
The paleobotanical site of Lava Camp Mine, on the northern side of the
Seward Peninsula only 75 km. south of the Arctic Circle, represents a
mid-Pliocene open wetland forest with many shaded pools all of which were
covered with a flow, of basaltic lava dated to 5.7 ± 0.2 m.y. The
preservation of actual woody specimens and numerous insects was excellent;
the flora represented consisted primarily of spruce and birch, but with
several other kinds of conifers and broad-leaved deciduous trees. The
combination of trees is not duplicated anywhere at present, but is more
reminiscent of southern Alaska and British Columbia than of today's western
Alaska. The numerous insects together indicate a semi-open forested environment
with some swamps and tree-shaded ponds, but not with dense forests, matched
best today by flood plains of the rivers of southern Alaska and British
Columbia. It was an environment of the type from which the Sasquatches
have often been reported and to which, if existing, sasquatches are well
adapted. Indeed, I suggest that that level of adaptation had already been
reached by the mid-Pliocene and that sasquatches at that time had already
reached Alaska.
During the late Pliocene and the early Pleistocene, the climate of the lands bordering Bering Sea became colder and Bering Isthmus became' a Bering Strait, as it is today. With the series of glacial periods that followed in the Pleistocene the environment became much more changeable, but even during the interglacial (such as the present) probably never again was as warm or as equable as it Gigantopithecus), these robust australopithecines have quite naturally been considered as possible ancestors of later giant hominoids. Additionally, on the basis of present evidence, they did not, make stone tools or know the use of fire, quite like Sasquatch, but neither did the other australopithecines or Gigantopithecus.
Possibly robust
australopithecines did extend their range beyond Africa and certainly
they could have evolved to become individuals of a size larger than we
know for Africa, where (in spite of exaggerated earlier claims) modern
research indicates a height varying around 5 ft., 4 inches (McHenry 1974).
The major problem of the robust australopithecines as possible ancestors
of Sasquatch, in my thinking, is their lateness in time; they are not
known earlier than 2,200,000 years ago, which figures admittedly may rely
only on lack of evidence, but even so I find difficulty in believing that,
if the animals ever did get out of Africa, they could have spread across
Asia into northeastern Siberia, and adapted to the Arctic cold typical
of that area by that time, and then crossed a Bering Bridge which was
exposed only under the super-Arctic conditions of a glacial period. I
find the idea of a Pleistocene robust australopithecine as ancestor for
Sasquatch much less convincing than that of a Pliocene Gigantopithecus.
C. Neanderthals have been recommended as ancestors for Eurasiatic "wildmen"
(Almas) and their North American kin (Sasquatches) by Heuvelmans and Porshnev
(1974), Porshnev (1974), and Bayanov and Bourtsev (1974). Note that I
did not, as most of my readers will think I should, label this Part C
as "Homo," since the scholars who recommend Neanderthals as
the prototype for the "wild-men" do not include them in Homo,
nor even in the Hominidae, whereas at least most American anthropologists
do so include them, as Homo sapiens neanderthalensis. The difference in
analysis of the available data is a profound one, due to one group (mostly
Russian) thinking in terms of the contrasting dichotomy of "animal"
and "man" whereas the other group (most Americans and British,
at least) think in terms of the similarities of "hominid animals" (including the australopithecines, Homo habilis. H. Erectus. H. Sapiens
neanderthalensis, and H. sapiens sapiens) and the differences, as
they exist, being primarily between such "hominid animals" and
the non-hominid animals (the remainder of the Animal Kingdom).
For reasons clearly outlined by Bayanov and Bourtsev (1974, 1976) those
who do not accept the human nature of the Neanderthals have established
a different family, the Troglodytidae, for neanderthalensis,
erectus, habilis, and the australopithecines, a family they regard
as not human but not apes, either. The basic distinction, I believe, is
that the troglodytes are regarded as not having abstract thinking
and speech whereas their "hominids" do. By separating modern
man from his nearer kin at the level of the family rather than that of
the subspecies one thinks of a considerable biological gulf between the
two groups, and the Troglodytidae; including the Neanderthals,
are automatically regarded as being much more primitive, primitive enough
that, if evolving larger size, they could become Almas and Sasquatches.
A considerable corpus of material culture (chipped stone tools, sometimes shelters, and usually remains of fires) are found associated with remains of Neanderthals and to a lesser degree with remains of some of the other "troglodytes."
In my mind, these
attributes of material culture are secondary energy traps and also tools
(Reed 1977, 1978), and their acquisition marked distinct steps toward
more utilization and control of the environment. Further, I regard the
Mousterian tool-kit, the mark of the Neanderthals over much of the Old
World, with its some 64 definite and repeated tool-types, to be evidence
for conceptual thinking involving time binding. I cannot agree that production
of such varied types of tools, seemingly to be used precisely for different
types of work, was "...a stereotyped and automatic function..."
and that it lay "...quite beyond the level of consciousness" (Porshnev 1974).
Additionally, I cannot agree that present evidence favors the supposition
that Neanderthals could not speak, or that even if they could produce
meaningful sounds they had no symbolic speech and thus no language. A
considerable history of denying speech or most aspects of intelligence
to Neanderthals has occurred, mostly on hypothetical preconceived bases
of imagined primitiveness at first and later on anatomical grounds (Lieberman
and Crelin, 1971; Lieberman, 1975), but at least in North America these
claims have been met by strong opposition, based on detailed research
by many anatomists. I find the arguments of the latter group (too numerous
to list here) to be convincing; additionally, the clear overall physical
resemblance between neanderthalensis and sapiens entitle them, on the
basis of comparison with populations of other species in nature, to no
more than subspecific differentiation. Further, there are the similarities
of the material culture (stone tools, shelters, use and presumably making
of fire, and probably clothing made of skins), which relate the two populations,
and additionally sapiens must have evolved from neanderthalensis; what
else was there? I am furthermore impressed with two other factors: 1)
The large brain of Neanderthals, larger on the average than is the brain
of some populations of living sapiens, must have been selected for problem-solving
at a level of complexity far above instinctual pattern-learning of the
details of an environment; the latter seems to be the level of selection
setting the intellectual level of the orangutan (Galdikas 1978). Neanderthals
buried their dead; even without the further level of complexity of burial
with grave-goods, the basic act of burial indicates to me a symbolic concept
of continuing existence after death; of that act by Neanderthals, Eiseley
(1957) has written, "We too were human, we too suffered, we too believed
that the grave is not the end. We too, whose face affright you now, knew
human agony, and human love." For the transmission of thoughts that
complex between members of the in-group and between individuals of different
generations, I regard symbolic speech to have been necessary.
On paleoenvironmental evidence, too, I must discount the Sasquatches--at least the North American Sasquatches--as modified Neanderthals; in spite of having a useful tool-kit, plus fire, shelter, and probably some clothing, Neanderthals never penetrated the high Arctic. They seemingly were never culturally adapted to a level of cold-resistant efficiency such that they could have survived in Siberia during the Pleistocene, not even in interglacial periods. They simply weren't there. If, with their Mousterian culture, a culture that allowed them to live immediately south of the European continental glacier during the period of Würm I, they could not penetrate the high Arctic, how could they be expected to do so if they abandoned that culture (fire, stone tools, shelter, and I think speech and clothing)? Certainly, the only route to North America for prehistoric man of the Pleistocene was across a Bering Bridge (Isthmus) or across the winter ice or summer waters of Bering Strait. Seemingly, Neanderthals never got into northeastern Siberia and I believe certainly never got into North America. I am convinced that that passage by hominids (using that term in North American terms) was accomplished in the late Pleistocene at least, only by anatomically modern man. Homo sapiens sapiens, the only Pleistocene hominid whose tools have been found in northeastern Siberia.
If and when we get a Sasquatch, living or dead, I foretell that it will not be a giant Neanderthal. In Eurasia, by contrast, there may well be two populations of "wildmen." Many of the depictions and descriptions there are of a size and morphology to be or have been surviving Neanderthals (Shackley 1982). Others, however, particularly in China, seem to me to be more similar in size and behavior to North American Sasquatches; if so, they are not Neanderthals (I say confidently) and may instead be descendants of the local populations of Chinese Gigantopithecus of half a million years ago.
Do we too, in North America, have two species of "wild-men", the true Sasquatches, and a population of more diminutive "apes" in the southern part of the United States?
© Charles Reed, 1984
References
Bayanov, Dmitri, and Igor Bourtsev
1974. Reply to B.F. Porshnev, The Troglodytidae and the Hominidae in the
Taxonomy and Evolution of Higher Primates. Current Anthropology 15 (4):452-456.
Bayanov, Dmitri, and Igor Bourtsev 1976.
On Neanderthal vs. Paranthropus. Current Anthropology 17 (2):312-318.
Eckhardt, R.B.
1975. Gigantopithecus As a Hominid. In Paleoanthropology,Morphology, and
Paleoecology. Russell H. Tutfie, ed. Pp. 105-129. The Hague: Mouton Publishers.
Eiseley, Loren
1957. Neanderthal Man and the Dawn of Human Paleontology.Quarterly Review
of Biology 32 (4):323-329.
Galdikas, Birute
1978. Orangutans and Hominid Evolution. In Spectrum: Essays Presented
to Sutan Takdir Alisjahbana on His Seventieth Birthday. S. Udin, ed. Pp.
287-309. Djakarta.
Heuvelmans, Bernard
and Boris Porshnev
1974. L'Homme de Neanderthal est Toujours Vivant. Paris: Plon.
Hopkins, D.M.
1967. The Cenozoic History of Beringia--A synthesis. In The Bering Land
Bridge. David M. Hopkins, ed. Pp. 451-454. Stanford: Standford University
Press.
Hopkins, D.M.J.V.
Matthews, J.A. Wolfe, and M.L. Silbersteing 1971. A
Pleiocene Flora and Insect Fauna From the Bering StraitRegion. paleogeography,
Paleoclimatology and Palaeo-
ecology 9 (3):211-231.
Lieberman, Philip
1975. On the Origins of Language: An Introduction To the Evolution of
Human Language. New York: Macmillan.
Lieberman, Philip,
and Edmund S. Crelin
1971. On the Speech of Neanderthal Man. Linguistic Inquiry 2 (2): 203-222.
McHenry, Henry
M.
1974. How Large Were the Australopithecines? American Journal of Physical
Anthropology 40 (3):329-340.
Petrov, O.M.
1967. Paleogeography of Chukotka During the Late Neocene and Quaternary
Times. In The Bering Land Bridge. David M.Hopkins, ed. Pp. 144-171. Stanford:
Stanford University Press.
Porshnev, B.F.
1974. The Troglodytidae and the Hominidae In the Taxonomy and Evolution
of Higher Primates. Current Anthropology15 (4):449-450.
Reed, Charles A.
1977. The Origins of Agriculture, Prologue. In Origins of Agriculture.
Charles A. Reed, ed. Pp. 9-21. The Hague: Mouton Press.
1978. Energy Traps, Tools and Culture. Abstracts of the Xth International Congress of Anthropological and Ethnological Sciences 2:253. Delhi, India, Dec. 10-21.
Rukang, Wu
1982a. Paleoanthropology In China, 1949-79. Current Anthropology 25 (5):473-477.
1982b. Recent Advances in Chinese Palaeoanthropology. Occasional Papers of the Hong Kong University Press 2:1-35.
Shackley, Myra
1982. The Case of Neanderthal Survival: Fact, Fiction or Faction? Antiquity
56 (216): 31-41.
Strasenburgh, Gordon
1975. On Paranthropus and "Relic Hominoids." Current Anthropology
16 (3):486-487.
1979. More on Neanderthal vs. Paranthropus. Current Anthropology 20 (3): 624-626.
Wolfe, Jack A.
1972. An Interpretation of Alaska Tertiary Floras. In Floristics and Paleofloristics
of Asia and Eastern North America. A. Graham, ed. Pp. 201-233. Amsterdam:
Elsevier Publishing.
Back to Biology, Scientific Papers and Field Reports
Back to What's New?
Back to Newspaper & Magazine Articles
Home/Main
Portions of this website are reprinted and sometimes edited to fit the standards of this website under the Fair Use Doctrine of International Copyright Law
as educational material without benefit of financial gain.
http://www4.law.cornell.edu/uscode/17/107.html
This proviso is applicable throughout the entire Bigfoot Encounters Website.